Klonieren
Ziel der molekularen Klonierungsverfahren ist es, ein gewünschtes Gen in einen geeigneten Expressionswirt einer kompatiblen Weise (rekombinante DNA) einzubringen, um schließlich das Genprodukt (Protein) in zufriedenstellender Qualität und Quantität herzustellen. Klonen bedeutet die Herstellung identischer Kopien eines bestimmten Gens von Interesse. Bei der Klonierung wird das Gen vervielfältigt und dann in ein Plasmid (Vektor) eingefügt, um es anschließend zu replizieren und das Protein zu exprimieren. Die Verwendung von Plasmiden ermöglicht den Schutz des fremden genetischen Materials vor den angeborenen Abbaumechanismen des Expressionswirts.

Es gibt verschiedene Klonierungsverfahren, um das gewünschte Gen in ein Plasmid einzufügen. Das Klonieren auf der Basis von Restriktionsenzymen ist die Standardklonierungsmethode in der Molekularbiologie. Restriktionsendonukleasen spalten doppelsträngige DNA (dsDNA) an spezifischen Sequenzstellen, den sogenannten Erkennungsstellen. Je nach verwendetem Restriktionsenzym können die erzeugten DNA-Fragmente entweder stumpfe Enden oder klebrige Enden haben. Diese können mit Plasmid-DNA fusioniert werden, wenn diese mit demselben Restriktionsenzym gespalten und dadurch linearisiert vorliegt. Die Fusionierung des DNA-Fragments und der linearisierten Plasmid-DNA erfolgt mit Hilfe einer sogenannten DNA-Ligase.
Eine spezielle Klonierungsmethode nutzt die spezifischen Eigenschaften von Typ-IIS-Restriktionsenzymen, Endonukleasen, die DNA außerhalb der Erkennungssequenz schneiden. Im Gegensatz zur traditionellen Klonierung mit Restriktionsenzymen bietet dieses Schneiden außerhalb der Erkennungsstelle den Vorteil, dass individuelle Überhänge erzeugt werden können. Diesen Ansatz hat IBA Lifesciences auch für das StarGate-System genutzt, um kompatible individuelle Überhänge zu erzeugen, die dann effizient fusioniert werden können. Das StarGate-System bietet eine große Anzahl an Klonierungs- und Expressionsvektoren für E. coli und Säugetierzellen.
StarGate für schnelles Klonieren
StarGate wurde für das schnelle systematische Screening des optimalen Expressions- (in Bezug auf den Expressionswirt und/oder den Promotor) und Reinigungssystems (in Bezug auf den Affinitäts-Tag) für ein bestimmtes Gen von Interesse (GOI) entwickelt.
Direktes Transfer-Klonieren

Das direkte Transferklonierungsverfahren wird empfohlen, wenn das optimale Expressions- und Reinigungssystem bereits bekannt ist. Bei diesem Verfahren kann ein PCR-Produkt, das das betreffende Gen enthält, direkt in den entsprechenden Expressionsvektor (Akzeptorvektor) eingefügt werden.
Im ersten Schritt wird das Gen von Interesse (GOI) an beiden Enden mit kombinatorischen Stellen (orange und rot) und den Esp3I-Erkennungsstellen (hellblau) über Polymerasekettenreaktion (PCR) ausgestattet, um eine Insertion in der korrekten Orientierung zu ermöglichen. Das PCR-Produkt wird im zweiten Schritt in den entsprechenden Expressionsvektor integriert, wodurch das endgültige Expressionskonstrukt, der so genannte Zielvektor, entsteht. Die Bildung des korrekten Zielvektors wird durch blau-weißes Screening auf LB-Agar-Platten mit X-Gal überprüft.
"Zwei-Schritt-Klonierung" über pENTRY-IBA
Die StarGate-Klonierung über den pENTRY-IBA-Vektor bietet ein Werkzeug für die systematische und schnelle Suche nach den besten Expressionsbedingungen für eine bestimmte GOI, für die in der Literatur nichts bekannt ist. Nach der anfänglichen pENTRY-Klonierung kann die GOI im resultierenden Donor-Vektor durch eine einfache Ein-Tube-Reaktion leicht in eine Vielzahl von Akzeptor-Vektoren mit unterschiedlichen Eigenschaften übertragen werden.

Schritt 1: Eingangsklonierung für die Erzeugung von Spendervektoren
Im ersten Schritt wird das Gen von Interesse (GOI) an beiden Enden mit kombinatorischen Stellen und den LguI-Erkennungsstellen ausgestattet, die für die orientierte Insertion des PCR-Fragments in pENTRY-IBA51 wichtig sind. Dies geschieht durch PCR unter Verwendung einer proofreading-Polymerase.
Die Rekombination des PCR-Produkts mit dem Eingangsvektor an den kombinatorischen Stellen (rot und orange) führt zur Erzeugung des Spendervektors. Bei diesem Schritt gehen die LguI-Restriktionsstellen (dunkelorange mit Pfeilspitzen) verloren, wodurch die Rekombinationsreaktion unidirektional und damit hocheffizient ist.
Im resultierenden Donor-Vektor werden dieselben kombinatorischen Stellen nun von den Esp3I-Erkennungsstellen (hellblau) flankiert, wodurch ein hocheffizienter und spezifischer StarGate®-Gentransferprozess in Akzeptor-Vektoren in ähnlicher Weise ermöglicht wird.
")
Schritt 2: Transfer-Klonierung (Erzeugung des Zielvektors)
In der Transferreaktion wird die GOI aus dem Donor-Vektor in einen Expressionsvektor (Akzeptor-Vektor) übertragen. Die Akzeptor-Vektoren bieten die gewünschte genetische Umgebung (d. h. Affinitäts-Tag, Promotor (Prom), zusätzliche Signalsequenzen usw.). In Fällen, in denen die optimalen Expressions- und Reinigungsbedingungen für die GOI nicht bekannt sind, sollten verschiedene Akzeptor-Vektoren ausgewählt werden.
Der Transfer vom Donor-Vektor in den Akzeptor-Vektor wird durch Esp3I als Restriktionsenzym und T4-DNA-Ligase vermittelt. Im resultierenden Zielvektor wird die GOI unter die Kontrolle des ausgewählten Promotors (Prom) gestellt, was die GOI-Expression im ausgewählten Expressionswirt ermöglicht. In diesem Beispiel ist ein Affinitäts-Tag (grün) an das C-terminale Ende des GOI-Expressionsprodukts fusioniert.
Besondere Merkmale der StarGate-Klonierung
Klonierung über Gensynthese
Gensynthese - der einfache Weg zur Erstellung Ihres StarGate-Donor-Vektors

So erzeugen Sie einen StarGate-Donor-Vektor mit Hilfe eines Gensynthesedienstes:
- Geben Sie die Sequenz Ihrer GOI ohne Start- und Stoppcodon ein
- Wählen Sie eine spezifische Codon-Optimierung für Ihren endgültigen Wirt. Fügen Sie die Esp3I-Erkennungsstelle und die entsprechenden kombinatorischen Stellen hinzu:
Start: 5'-AGCGCGTCTCCAATG
Stopp: 3'-GGGAGGAGACGCGCT - Wählen Sie einen Standardklonierungsvektor (vom Dienstleister angeboten), der ein Antibiotikaresistenzgen für Kanamycin enthält.
Expressionsvektoren (Akzeptor-Vektor)
IBA-Akzeptorvektoren bieten verschiedene genetische Umgebungen für eine optimale Proteinexpression und -reinigung. Dazu gehören verschiedene Reinigungs-Tags, Promotoren für eine Vielzahl von Wirtszellen, Signalsequenzen usw.
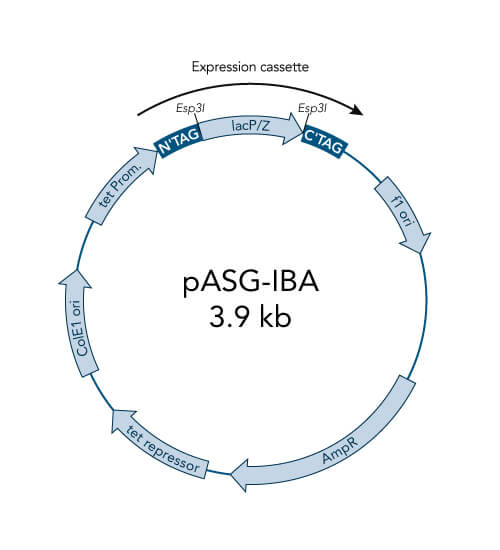
E.Coli
pASG-IBA vectors
- High-level expression in E. coli, also for toxic proteins
- Tightly regulated expression due to anhydrotetracycline (AHT) inducible tetA promoter/operator
- Option for periplasmic expression due to ompA signal sequence
- No catabolite repression - no influence of medium components
- Not influenced by the genetic background – wide choice of E. coli expression strains
- Inexpensive induction with AHT
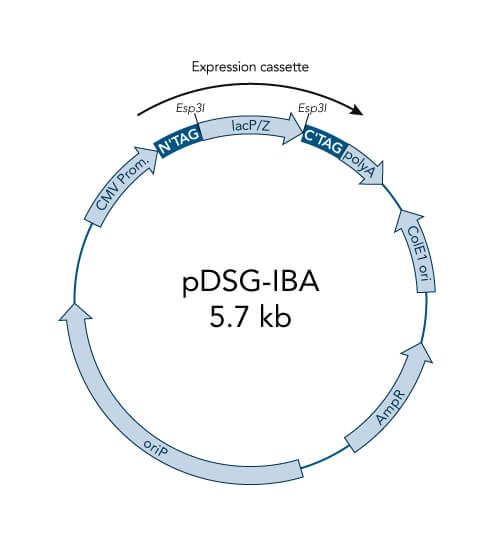
Mammalia
pDSG-IBA vectors
- Optimized for MEXi, IBA´s mammalian expression system, and are also suitable for other mammalian cell cultures
- High level constitutive expression in mammalian cells by the CMV promoter
- Extrachromosomal replication due to Epstein Barr Virus replication origin (oriP) - requires chromosomal expression of the EBNA-1 gene, e.g. like in MEXi 293E cells
- ampicillin resistance cassette for selection of transformed E. coli cells
- ColE1 origin for a high plasmid copy number
- BM40 option for secretion of protein into the medium
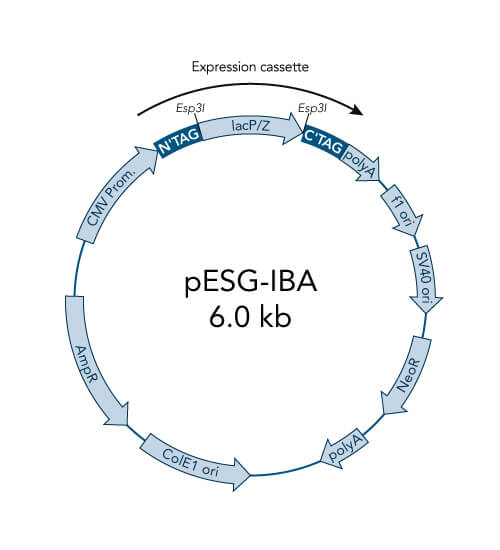
Mammalia
pESG-IBA vectors
- High-level constitutive expression in mammalian cells by the CMV promoter
- Neomycin resistance for generation of stable cell lines
- ampicillin resistance cassette for selection of transformed E. coli cells
- ColE1 origin for a high plasmid copy number
- BM40 option for secretion of protein into the medium
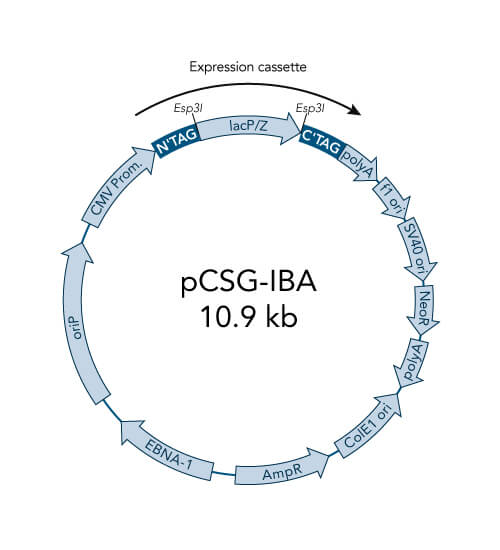
Mammalia
pCSG-IBA vectors
- High-level constitutive expression in mammalian cells by the CMV promoter
- extrachromosomal replication due to Epstein Barr Virus replication origin (oriP) and
- chromosomal expression of the EBNA-1 gene is not required, because the EBNA-1 is encoded by the vector
- prolonged expression of the inserted GOI under G418 selection without the need for making stable cell lines
- ampicillin resistance cassette for selection of transformed E. coli cells
- ColE1 origin for a high plasmid copy number
- BM40 option for secretion of protein into the medium
Tet-Expressionssystem

Merkmale und Vorteile der pASK-IBA-Vektoren:
- Hochgradige Expression in E. coli auch für zytotoxische Proteine
- Streng regulierte Expression durch den Tetracyclin-Promotor
- Kostengünstige Induktion mit Anhydrotetracyclin
Die pASK-IBA-Vektoren sind mit Strep-tag®II, dem OmpA-Sekretionssignal, speziellen Proteaseschnittstellen und Chloramphenicol-Resistenz erhältlich.
Prinzip und Eigenschaften
pASK-IBA-Vektoren enthalten den streng regulierten Tetracyclin (tetA)-Promotor, der bis zur Induktion mit einer niedrigen Konzentration an Anhydrotetracyclin durch den tet-Repressor unterdrückt wird. Eine vorzeitige Expression des fremden Gens bleibt dadurch aus. Der tet-Repressor ist ebenfalls auf dem pASK-IBA-Vektoren kodiert und wird konstitutiv vom Chloramphenicol-Acetyltransferase-Promotor exprimiert. Die Vektoren vermitteln aber keine Resistenz gegen Tetracyclin.
Anders als beim T7-Promotor sind keine speziellen E. coli-Stämme oder zusätzliche Plasmide erforderlich. Im Gegensatz zum lac-Promotor liegt beim tetA-Promotor/Operator keine vorzeitige Expression ohne Induktionsmittel vor und ist funktionell nicht an zelluläre Regulationsmechanismen oder einen bestimmten genetischen Hintergrund gekoppelt. Des Weitere ist der tetA-Promotor im Vergleich zum lac-Promotor auch nicht anfällig für Katabolitrepression (cAMP-Spiegel, Stoffwechselzustand), wird nicht von chromosomal kodierten Repressormolekülen beeinflusst.
Infolgedessen sind keine speziellen E. coli-Stämme oder zusätzliche Plasmide erforderlich, und es kann eine breite Palette von Kulturmedien und -bedingungen verwendet werden. So können beispielsweise Glukose-Minimalmedien und sogar der Bakterienstamm XL1-Blue, der eine episomale Kopie des Tetracyclin-Resistenzgens trägt, zur Expression verwendet werden. Das pASK-IBA-Expressionssystem ist unter vielen Bedingungen, einschließlich Fermentation, stabil und einfach zu handhaben.
Weitere Elemente der Vektoren sind eine Tandem-Ribosomenbindungsstelle (RBS), die eine effiziente Initiierung der Translation gewährleistet, der starke Terminator des Lipoprotein-Gens, um ein Durchlesen zu verhindern, die intergene Region des Bakteriophagen f1, die eine Möglichkeit zur Herstellung von ssDNA bietet, sowie ein Chloramphenicol-Acetyltransferase-Gen.
Referenz: Skerra, A. (1994). Use of the tetracycline promoter for the tightly regulated production of a murine antibody fragment in Escherichia coli. Gene, 151, 131-135.
Die folgenden E. coli-Stämme wurden bereits erfolgreich für die Tet-Expression mit unseren pASK-IBA-Vektoren verwendet:
- JM83
- WK6
- B
- BL21
- MG1655
- W3110
- BL21(DE3)
- BLR(DE3)
- XL1-Blue
- BL21-CodonPlusTM-RIL
Für die Sekretion empfehlen wir JM83. Für die zytoplasmatische Expression werden E. coli B-Stämme empfohlen, da ihnen die lon-Protease und die ompT-Protease der äußeren Membran fehlen, die Proteine abbauen können (Grodberg und Dunn, 1988, J.Bacteriol. 170, 1245).
Bisher ist uns kein E. coli-Stamm bekannt, der mit dem Tet-Expressionssystem inkompatibel ist.
Anhydrotetracyclin: Induktor für den tetA-Promotor
Die Expressionskassette der pASK-Vektoren steht unter der Transkriptionskontrolle des tetA-Promotors/Operators und Repressors. Der Promotor wird durch eine niedrige Konzentration von Anhydrotetracyclin (AHT) induziert, was Kosten spart und die antibakterielle Wirkung von AHT minimiert. Degenkolb et al. (1991) haben gezeigt, dass AHT 35-mal fester als Tetracyclin an den tet-Repressor bindet.
TypIIS-Restriktionsenzyme
Typ-II-Enzyme sind eine der vier (I-IV) Arten von anerkannten Endonukleasen, die DNA an einer bestimmten Erkennungsstelle schneiden. Typ-II-Enzyme schneiden innerhalb oder in geringer Entfernung von ihren Erkennungsstellen, die in der Regel 4-8 Nukleotide lang sind.
Zu ihnen gehören die Enzyme vom Typ IIS, wie LguI und Esp3I.
Restriktionsenzyme vom Typ IIS sind dimere Enzyme, die DNA in einem bestimmten Abstand von ihrer nicht-palindromischen, asymmetrischen Erkennungsstelle spalten. Dies bedeutet, dass die Zielsequenz nur in einer Richtung gelesen werden kann. Dadurch kann der Verdau mit nur einem einzigen Enzym zwei verschiedene unabhängige klebrige Enden mit 5'-Überhängen erzeugen, die eine gerichtete Klonierung ermöglichen. Außerdem wird nach der Verdauungsreaktion die Erkennungssequenz vollständig entfernt, so dass die kodierte Aminosäuresequenz durch die verbleibenden Restriktionsenzymstellen nicht beeinträchtigt wird.

Die Verwendung von Restriktionsenzymen vom TypIIS bietet wichtige Funktionen für die Klonierung:
- Sie ermöglicht das Klonieren in einem Reaktionsgefäß
- Die exprimierten Proteine enthalten keine zusätzlichen Aminosäuren
- Die Klonierung erfolgt immer im gleichen Leserahmen der weiteren Vektoreigenschaften
- Der Zusammenbau mehrerer Fragmente ist möglich
Referenzen: Pingoud A, Jeltsch A (2001). Structure and function of type II restriction endonucleases. Nucleic Acids Res. 29 (18): 3705–27.
Anwendungsbeispiel
StarGate umfasst Vektoren zur Expression in Säugetierzellen (pCSG-IBA, pESG-IBA, pDSG-IBA) und E. coli (pASG-IBA). Als Beispiel wurde das bakterielle Protein Azurin in verschiedene pASG-IBA-Vektoren kloniert und in E. coli exprimiert. Die Expressionsergebnisse wurden mittels SDS-PAGE analysiert.

Die kodierende Sequenz von Azurin, einem 14-kDa-großen bakteriellen Protein, wurde in 9 verschiedene pASG-IBA-Vektoren kloniert und in E. coli exprimiert. Vergleichbare Mengen von E. coli-Zellen wurden 3 Stunden nach der Induktion mit Anhydrotetracyclin geerntet. Die Proben wurden durch 5-minütiges Kochen bei 95 °C mit Gelladepuffer lysiert und mittels SDS-PAGE analysiert. Anschließend wurde das Gel mit Coomassie angefärbt. Die periplasmatische Sekretion mittels ompA führte in allen Fällen zur Anreicherung vergleichbarer Mengen des Zielproteins (Spuren 6-9). Dies war zu erwarten, da Azurin auch unter natürlichen Bedingungen in P. aeruginosa sezerniert wird. Im Falle der zytosolischen Expression zeigten sich jedoch überraschend, dass eine zytosolische Expression von Azurin überhaupt möglich ist. Die Expression wurde durch die Fusion eines N-terminalen Affinitäts-Tags ermöglicht (Spur 2, 4 und 5), während N-terminal nicht getaggte Varianten zu keiner Expression führten (Spur 1 und 3). Das Beispiel zeigt, dass sich ein anfängliches Screening der Expressionsbedingungen lohnen kann, da verschiedene Konstrukte je nach den Eigenschaften des Zielproteins zu unterschiedlichen Ergebnissen führen können.
